


糖鎖の生合成を糖鎖自身が制御する仕組みを発見
2024年9月30日
岐阜大学
糖鎖の生合成を糖鎖自身が制御する仕組みを発見
【本研究のポイント】
・タンパク質に付いた糖鎖に枝分かれを作る酵素GnT-IVaとGnT-IVbは、特徴的なレクチンドメイン(糖鎖結合領域)を持つ
・GnT-IVaと-IVbのレクチンドメインは糖鎖を付ける相手のタンパク質を見分ける
・GnT-IVaと-IVb自身にも糖鎖が付いており、自分自身の糖鎖がレクチンドメインに結合する。この結合によって酵素の糖鎖枝分かれを作る働きが阻害される
・特定の構造の糖鎖が、同じ構造の糖鎖の合成を抑制するという、自己制御の仕組みを発見した
【研究概要】
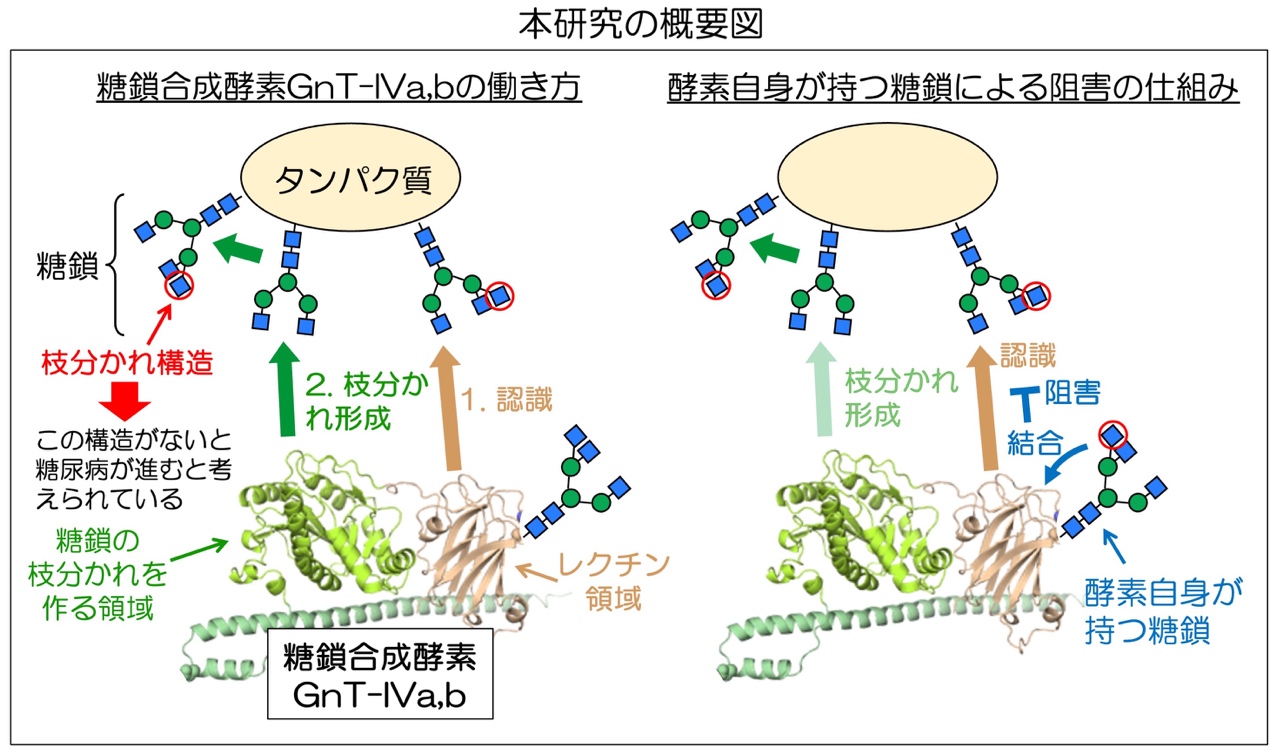
岐阜大学糖鎖生命コア研究所の木塚 康彦教授、自然科学技術研究科修了生の長田菜緒子さんらの研究グループは、ミシシッピ大学、広島大学、大阪大学との共同研究で、タンパク質に付いた糖鎖の枝分かれ構造が、この枝分かれを作る酵素の働きを抑える、つまり特定の糖鎖が自分自身の生合成を抑えるという新たな仕組みを発見しました。
タンパク質に付く糖鎖には膨大な種類があり、細胞の中で多くの糖鎖合成酵素により作られます。それぞれの糖鎖の量はこれら酵素の働きにより正しく制御され、それが疾患により異常となることが報告されていますが、酵素の働きを制御する仕組みはよくわかっていません。
GnT-IVaとGnT-IVbはそれら糖鎖合成酵素の一つで、糖鎖の枝分かれを作ります。GnT-IVaを欠損するマウスは糖尿病様の症状を示すことから、GnT-IVaは糖尿病との関連が示されています。また、GnT-IVaと-IVbは、他の糖鎖合成酵素が持たないレクチンドメイン(糖鎖結合領域)を分子内に持っていますが、その役割は不明でした。本研究では、このレクチンドメインが、糖鎖を持つ基質タンパク質の認識に必要であることを明らかにしました。さらに、GnT-IVaと-IVb自身にも糖鎖が付いており、この酵素自身の糖鎖が枝分かれ構造を持つと、レクチンドメインに結合してその機能を阻害し、GnT-IVaと-IVbによる枝分かれ形成を低下させることがわかりました。
本研究は、特定の糖鎖が自分自身の合成を阻害するという、糖鎖による自己抑制の仕組みを初めて示しました。以上の成果は、複雑な糖鎖形成の仕組みの解明と、糖尿病の病態解明や治療薬開発への応用が期待されます。
本研究成果は、現地時間2024年9月27日付で『iScience』に掲載されました。また本研究は、文部科学省の大規模学術フロンティア促進事業「ヒューマングライコームプロジェクト」により支援を受けました。
【研究背景】
糖鎖1)とは、グルコースなどの糖(動物では約10種類の糖が存在)が枝分かれしながら鎖状につながったもので、多くはタンパク質や脂質などに結合した状態で存在しています。動物では、体内の半数程度のタンパク質に糖鎖が付いていると考えられており、糖鎖を持つタンパク質は糖タンパク質と呼ばれています。タンパク質に付いている糖鎖には様々な形のものがあり、タンパク質ごとに形が異なることや、同じタンパク質でも、健康なときと病気のときとで糖鎖の形が変化することが知られています。こうした疾患特異的な糖鎖の変化は、実際に医療の現場でがんの診断などに使われています。また、特定の糖鎖が、がんやアルツハイマー病など様々な疾患の発症や進行に重要な役割を果たすことから、糖鎖を標的とした新たな治療薬の開発が期待されています。
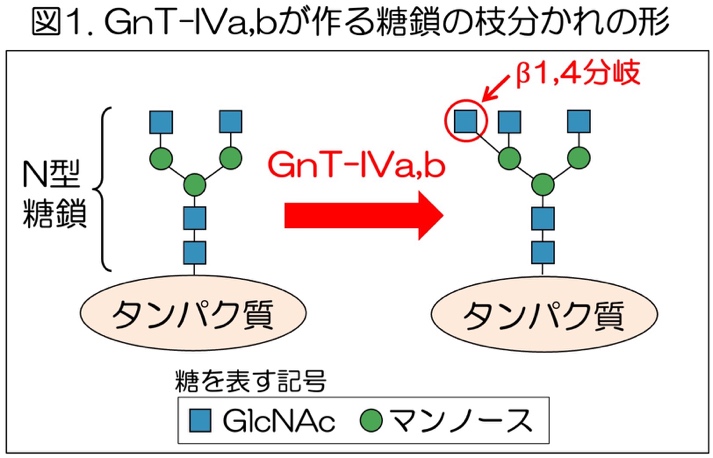
タンパク質に付く糖鎖は、細胞の中で糖転移酵素2)(糖鎖合成酵素)と呼ばれる酵素の働きによって作られます。ヒトの体内には、約180種類ほどの糖転移酵素が存在し、それらの働きが厳密に制御されることで膨大な種類の糖鎖構造を作ります。しかし、個々の酵素の働きを制御する仕組みはよくわかっておらず、生物がどのように膨大な種類の糖鎖の量を調節しているかはまだあまりよくわかっていません。糖転移酵素のうち、GnT-IVaとGnT-IVb 3)は、タンパク質に付いたN型糖鎖4)と呼ばれる糖鎖に作用し、糖鎖の枝分かれ構造を作ります(図1)。これまでの研究で、GnT-IVaは血糖調節に重要な膵臓に多く存在し、GnT-IVaを欠損させたマウスは高血糖やインスリン分泌不全などの糖尿病様の症状を示すことから、GnT-IVaが作る糖鎖は糖尿病の発症・進行と関係があることがわかっています。
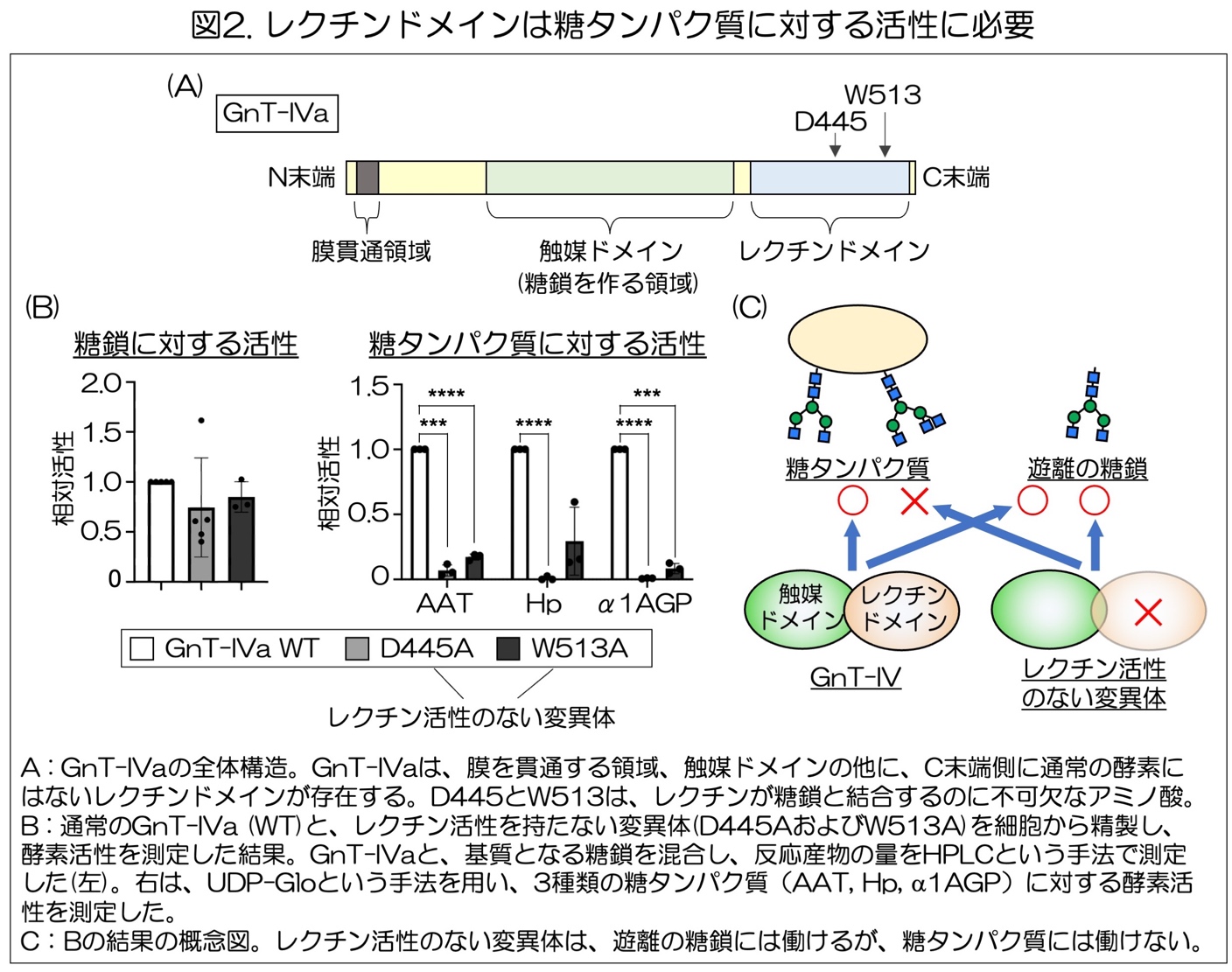
このGnT-IVaとIVbは、他の糖転移酵素とは異なる構造を持っており、糖鎖を作る触媒5)ドメイン6)の他に、特定の糖鎖と結合するレクチン7)ドメインを持っていることが、木塚教授らの最近の研究でわかってきました(図2A)(https://igcore.thers.ac.jp/wp_control/wp-content/uploads/79e08b70c6b99f0115ad4429a1b80905-2.pdf)。しかし、このレクチンドメインが持つ役割についてはよくわかっていませんでした。
【研究成果】
本研究ではまず、GnT-IVaとIVbのレクチンドメインの役割を明らかにするために、レクチンドメインが糖鎖と結合するのに不可欠な2つのアミノ酸(D445、W513)を変異させ、レクチンドメインが糖鎖と結合できないGnT-IVaの変異体8)を作りました(図2A)。この変異体を用いて酵素の活性を測定したところ、遊離のN型糖鎖に対する枝分かれ活性は、通常のGnT-IVa (WT)とレクチン活性のない変異体との間で大きな変化はありませんでしたが(図2B左)、N型糖鎖を持つ糖タンパク質に対する活性は、これらの変異体で劇的に低下していました(図2B右、図2C)。このことから、GnT-IVaやIVbのレクチンドメインは、糖タンパク質に対する酵素活性に不可欠であり、枝分かれ構造を作るべき基質9)糖タンパク質を認識する働きがあることが示されました。
また、GnT-IVaやIVbは、それら自身も糖タンパク質であり、糖鎖が付いています。
GnT-IVaには2箇所のN型糖鎖が付いており、1つはレクチンドメインに付いています(N449に付いた糖鎖)(図3A)。さらにこの糖鎖は、レクチンドメインの糖鎖認識部位(図3A, *)と極めて近い位置にあります。そこで木塚教授らは、N449に付いた酵素自身のN型糖鎖が、レクチンドメインの糖鎖認識部位と結合し、レクチンドメインの機能を阻害するのではないかと仮説を立てました。
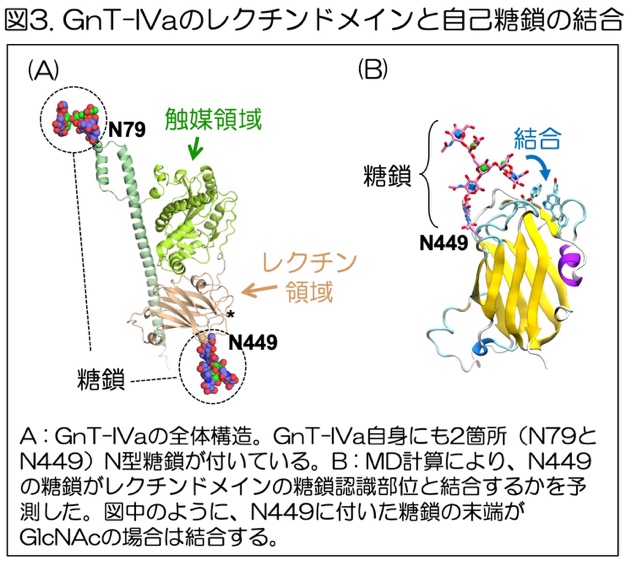
実際に、レクチンドメインが酵素自身の糖鎖と結合しているかどうかを明らかにするために、分子動力学シミュレーション10)という方法を使いました。その結果、N449に付いた糖鎖は、糖鎖の構造に依存してレクチンドメインの糖鎖認識部位と結合することがわかりました(図3B)。特に、N449に付いた糖鎖の形が、GnT-IVaやIVbが作る枝分かれ構造を持ったN型糖鎖であるとき、N449に付いた糖鎖とレクチンドメインとの結合が強いことがわかりました。以上から、GnT-IVaと-IVb自身に付いたN型糖鎖は、レクチンドメインに結合して機能を抑える、自己リガンド11)として働きうることがわかりました。
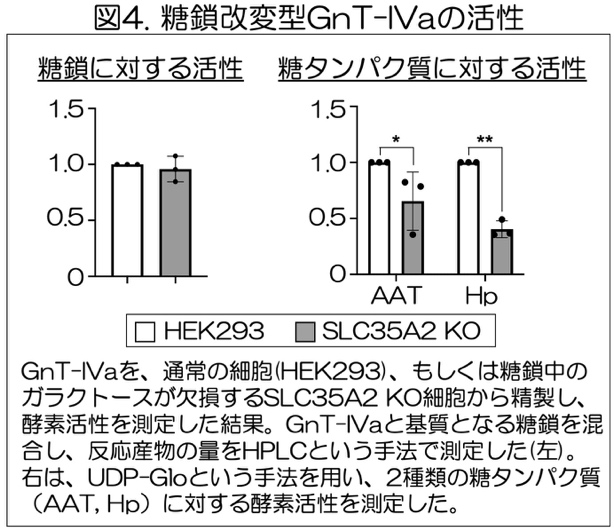
最後に、この酵素自身の糖鎖のレクチンドメインへの結合によって、酵素活性が阻害されるかどうかを調べました。そのために、レクチンドメインのリガンドとならない形の糖鎖をもったGnT-IVaと、リガンドとなる形の糖鎖をもったGnT-IVaを、それぞれHEK293細胞、SLC35A2-KO HEK293細胞から精製することで調製し、活性を測定しました。その結果、遊離のN型糖鎖に対する活性は、自分自身がリガンド糖鎖をもっていてもいなくても変わらない(図4左)のに対し、糖タンパク質に対する活性は、レクチンドメインのリガンドとなる糖鎖をもったGnT-IVa(SLC35A2KO細胞由来)の方が低いことがわかりました(図4右)。以上より、酵素自身の糖鎖がレクチンドメインと結合すると、レクチンドメインによる基質タンパク質の認識が妨げられ、糖タンパク質への枝分かれ形成が抑えられることが明らかになりました。
【今後の展開】
本研究により、GnT-IVa, IVbは、ユニークなレクチンドメインを介して、作用する基質タンパク質を認識していること、さらに、酵素自身の糖鎖によってその認識を阻害し、活性を抑えていることがわかりました。
本研究は、ヒトが持つ多様な糖鎖の複雑な生合成の仕組みの一端を明らかにしました。本研究は今後、糖鎖が作られる過程の全容解明や、糖鎖の生合成を自在に操る技術開発、糖尿病の病態メカニズムの理解および治療薬開発などへの応用が期待されます。
【論文情報】
雑誌名:iScience
タイトル:Self-regulation of MGAT4A and MGAT4B activity toward glycoproteins through interaction of lectin domain with their own N-glycans
著者:Naoko Osada, Sushil K. Mishra, Miyako Nakano, Yuko Tokoro, Masamichi Nagae, Robert J. Doerksen and Yasuhiko Kizuka
DOI番号: 10.1016/j.isci.2024.111066
論文公開URL:https://www.cell.com/iscience/fulltext/S2589-0042(24)02291-0
【用語解説】
1)糖鎖:グルコース(ブドウ糖)などの糖が鎖状につながった物質。遊離の状態で存在するものもあれば、タンパク質や脂質に結合した状態のものもある。デンプン、グリコーゲンなどの多糖は数多くの糖がつながり、糖鎖だけで遊離の状態で存在する。一方タンパク質に結合したものは、数個から20個程度の糖がつながったものが多い。
2)糖転移酵素:糖鎖を合成する酵素のことで、ヒトでは180種類程度存在することが知られている。主に、細胞の中のゴルジ体と呼ばれる小器官に存在している。
3)GnT-IVa, GnT-IVb:糖鎖を合成する糖転移酵素の一種で、細胞の中のゴルジ体に存在し、β1,4分岐という糖鎖の枝分かれ構造を作る。GnT-IVaとIVbはアミノ酸配列の類似性が高く、性質が極めてよく似ている。
4)N型糖鎖:タンパク質に付く糖鎖の種類の1つで、タンパク質のアスパラギン残基(アミノ酸の1文字表記でN)に結合している。ヒトでは7,000種類以上のタンパク質がN型糖鎖を持つと考えられている。
5)触媒:化学反応の速度を高める物質のこと。触媒自身は反応前後では変化しない。酵素は生体内の様々な反応を触媒するタンパク質である。
6)ドメイン:タンパク質の構造の一部のうち、他の部分とは独立して折り畳まれた領域のこと。一般に、タンパク質は複数のドメインからなる。
7)レクチン:様々な糖鎖や糖と選択的に結合するタンパク質の総称。糖鎖と結合するタンパク質であっても、抗体はレクチンに含まれない。ヒトの体内のレクチンは、先天性免疫などに重要な役割を果たす。
8)変異体:遺伝子配列に変異が入った個体や、変異が入った遺伝子から生じたタンパク質のことを指す。タンパク質の場合は、元になる遺伝子配列の変異によって、タンパク質のアミノ酸配列の一部が、通常とは異なる配列になっている。
9)基質:化学反応における出発物質のこと。反対に、化学反応の結果できるものを生成物(産物)と言う。酵素は、自身が触媒する反応において、特定の構造を持った基質を厳密に認識することから、基質と酵素は鍵と鍵穴の関係に例えられる。
10)分子動力学シミュレーション:Molecular Dynamics (MD)計算とも呼ばれ、分子や原子の動きを計算によって予測する方法。
11)リガンド:タンパク質などの生体分子と結合して複合体を形成する物質のこと。この場合は、レクチンに結合する糖鎖のこと。
.jpg)
-1.jpg)
_クレジット追加.jpg)
.jpg)




